Нуклеотиды в питании детей раннего возраста. Словарь Азотистое основание в составе нуклеотида атф
Наряду с аминокислотами важнейшей группой азотистых веществ являются нуклеотиды. Их биологическое значение для жизнедеятельности организмов определяется тем, что они используются для построения молекул нуклеиновых кислот - дезоксирибонуклеиновой (ДНК) и рибонук-леиновой (РНК), входят в состав каталитических центров ферментов, участвуют в биоэнергетических процессах и синтезе углеводов, липидов, белков, алкалоидов и других веществ. Некоторые нуклеотиды способны выполнять регуляторные функции.
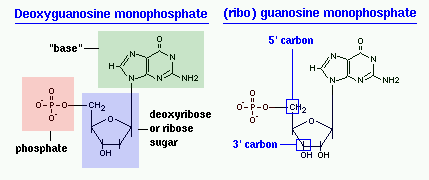
Главные структурные компоненты нуклеотидов–азотистые основания, пентозы (рибоза или дезоксирибоза) и остаток ортофосфорной кислоты. В зависимости от углеводного компонента различают две группы нук-леотидов: рибонуклеотиды, содержащие остаток рибозы, и дезоксирибо-нуклеотиды, имеющие в своем составе остаток дезоксирибозы. Дезоксирибонуклеотиды используются организмами для синтеза ДНК, а рибонуклетиды входят в состав РНК, ферментов и макроэргических нук-леозидполифосфатов.
Рибоза и дезоксирибоза в составе нуклеотидов находятся в b-D-фура-нозной форме:
Нуклеотиды образуются из двух типов азотистых оснований – произ-водных пиримидина и пурина. Свойства оснований они проявляют в водном растворе при взаимодействии с молекулами воды. Из пиримидиновых осно-ваний наиболее важное значение имеют урацил, тимин и цитозин как основные структурные единицы нуклеотидов, образующих нуклеиновые кислоты. Кроме них, известны и другие основания - 5-метилцитозин, псевдоурацил, 5-оксиметилцитозин и др. 5-Метилцитозин и 5-оксиметилцитозин в небольшом количестве могут
Из пуриновых оснований наибольшее значение имеют аденин и гуанин, так как они используются для синтеза нуклеиновых кислот. В составе нуклеиновых кислот в небольшом количестве обнаружены также и другие основания, которые образуются в результате химической модификации аденина и гуанина: 7-метилгуанин, 2-метиладенин, N-диметилгуанин и др. Важными промежуточными метаболитами являются гипоксантин, ксантин, аллантоин. В некоторых растениях они могут накапливаться в свободном состоянии.
Все азотистые основания интенсивно поглощают ультрафиолетовый свет при длинах волн 200-280нм.
При соединении азотистых оснований с молекулой рибозы или дезоксирибозы образуются соединения, называемые нуклеозидами , так как между пентозой и основанием возникает гликозидная связь. Основания в данном случае можно рассматривать как агликоны по отношению к пентозе.
В нуклеозидах гликозидная связь возникает между первым углеродным атомом пентозы в b-фуранозной форме и азотом пуринового (в девятом положении) или пиримидинового (в первом положении) основания. Азотистые основания аденин, гуанин, цитозин и урацил образуют при со-единении с рибозой нуклеозиды - аденозин, гуанозин, цитидин и уридин,
а с дезоксирибозой – дезоксиаденозин, дезоксигуанозин, дезоксицитидин, дезоксиуридин. Тимин, соединяясь с дезоксирибозой, даёт дезоксити-мидин.
Азотистые основания и нуклеозиды могут накапливаться в растениях в значительном количестве при интенсивном распаде нуклеиновых кислот.
Фосфорнокислые эфиры нуклеозидов называют нуклеотидами . В составе нуклеотидов остатки ортофосфорной кислоты могут присоединяться к пятому или третьему атомам углерода рибозы или дезоксирибозы, а у некоторых рибонуклеотидов ещё и ко второму атому углерода рибозы. У свободных нуклеотидов фосфатная группа обычно находится у пятого углеродного атома рибозы или дезоксирибозы. В нейтральной среде остатки ортофосфорной кислоты в молекулах нуклеотидов сильно диссоциированы, вследствие чего могут присоединять катионы, поэтому при химическом выделении нуклеотиды кристаллизуются в виде солей.
Изучение пространственной структуры азотистых оснований методом рентгеноструктурного анализа показывает, что все они имеют почти плоскую конформацию. У них довольно легко происходит перегруппировка двойных связей, которая сопровождается таутомерными превращениями. Например, гуанин может существовать в виде двух таутомерных форм:
Плоскость гетероциклического ядра основания в структуре нуклеозидов и нуклеотидов может занимать в пространстве два положения по отношению к пентозе, образуя две противоположные конформации - син -конформацию и анти -конформацию. В анти -конформации структура азотистого основания развернута от пентозы, а в син -конформации ориентирована над её плоскостью. В свободном состоянии пиримидиновые нуклеотиды находятся преимущественно в анти -конформации, а пуриновые довольно легко переходят из одной формы в другую.
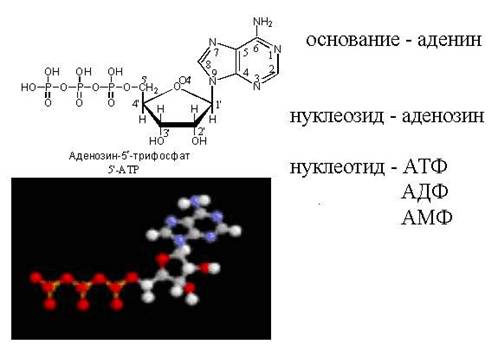
В связи с тем, что у нуклеотидов сильно выражены кислотные свойства, их называют кислотами с учетом названий азотистых оснований и углеводного компонента. Так, например, рибонуклеотид, имеющий остаток аденина, называют адениловой кислотой, или аденозинмонофосфатом (АМФ). Дезоксирибонуклеотид, образованный из тимина, называют дезокситимидиловой кислотой, или дезокситимидинмонофосфатом (дТМФ). Названия других нуклеотидов представлены в таблице 2.
В растениях найдены циклические формы нуклеотидов – адено-зинмонофосфата и гуанозинмонофосфата, которые по-видимому выполняют регуляторные функции. Строение циклического АМФ можно представить слудующей формулой:
2 . Названия важнейших нуклеотидов.
Нуклеотид – нуклеозид + один или несколько остатков фосфорной кислоты. Нуклеозид – азотистое основание и молекула пентозы. В состав нуклеотидов входят два пуриновых основания (аденин и гуанин) и 3 пиримидиновых основания (тимин, урацил, цитозин). Иногда встречаются минорные азотистые основания: псевдоурацил, метилуридин, метилцитозин, метиладенин.
Номенклатура:
Первичная структура НК – полинуклеотидная цепь со строго определенной последовательностью нуклеотидов, соединенных между собой 3’-5’-фосфодиэфирной связью.
Свойства нуклеотидов : 1) приобретают отрицательный заряд 2) обладают ярко
Выраженными кислотными свойствами.
Особенности строения, функции и распределения в клетке ДНК и РНК:
|
Локализована в основном в ядре, также в митохондриях и хлоропластах |
Локализована в основном в цитоплазме |
|
В структуру входят А, Т, Г, Ц + дезоксирибоза + остаток фосфорной к-ты. |
В структуру входят А, У, Г, Ц + рибоза + остаток фосфорной к-ты |
|
Двойная спираль (известно 6 типов: А-Е, Z, преобладающая B-форма) |
Одноцепочечная (хотя и может сворачиваться с образованием «шпилек»). Имеет разновидности (иРНК, мРНК, тРНК) |
|
Различаются по размерам (ДНК обычно состоит из большого количества нуклеотидов) |
|
|
1. Обеспечивает синтез белка 2. Носитель наследственной информации |
Обеспечивают синтез белка |
|
Подчиняется правилам Чаргаффа |
Не подчиняется правилам Чаргаффа |
Метод анализа первичной структуры ДНК (Сэнджер):
Основан на ДНК-полимеразной реакции: выделение ДНК ® разрезание ее рестриктазами ® денатурация фрагментов ДНК и получение одноцепочечных молекул, используемых в качестве матрицы ® добавляют праймер и субстраты для синтеза ДНК ® смесь делят на четыре пробирки, в каждую добавляют один из стоп-нуклеотидов (дидезоксинуклеотидов) и ДНК-полимеразу ® синтез останавливается при встрече ДНК-полимеразой стоп-нуклеотида ® после окончания в каждой пробирке находятся фрагменты, заканчивающиеся на определенный нуклеотидов ® фрагменты разделяют электрофорезом в агарозном геле и анализируют.
- это сложные мономеры, из которых собраны гетерополимерные молекулы. ДНК и РНК. Свободные нуклеотиды участвуют в сигнальных и энергетических процессах жизнедеятельности. ДНК-нуклеотиды и РНК-нуклеотиды имеют общий план строения, но различаются по строению сахара-пентозы. В ДНК-нуклеотидах используется сахар дезоксирибоза, а в РНК-нуклеотидах - рибоза.
Структура нуклеотида
В каждом нуклеотиде можно выделить 3 части:
1. Углевод - это пятичленный сахар-пентоза (рибоза или дезоксирибоза).
2. Фосфорный остаток (фосфат) - это остаток фосфорной кислоты.
3. Азотистое основание - это соединение, в котором много атомов азота. В нуклеиновых кислотах используется всего 5 видов азотистых оснований: Аденин, Тимин, Гуанин, Цитозин, Урацил. В ДНК - 4 вида: Аденин, Тимин, Гуанин, Цитозин. В РНК - тоже 4 вида: Аденин, Урацил, Гуанин, Цитозин, Легко заметить, что в РНК происходит замещение Тимина на Урацил по сравнению с ДНК.
Общая структурная формула пентозы (рибозы или дезоксирибозы), молекулы которой образуют "скелет" нуклеиновых кислот:
Если Х заменить на Н (Х = Н) - то получаются дезоксирибонуклеозиды; если Х заменить на ОН (Х = ОН) - то получаются рибонуклеозиды. Если вместо R подставить азотистое основание (пуриновое или пиримидиновое) - то получится конкретный нуклеотид.
Важно обратить внимание на те положения атомов углерода в пентозе, которые обозначены как 3" и 5". Нумерация атомов углерода начинается от атома кислорода вверху и идёт по часовой стрелке. Последним получается атом углерода (5"), который располагается за пределами пентозного кольца и образует, можно сказать, "хвостик" у пентозы. Так вот, при наращивании цепочки из нуклеотидов фермент может присоединить новый нуклеотид только к углероду 3" и ни к какому другому. Поэтому 5"-конец нуклеотидной цепочки никогда не сможет иметь продолжения, удлинняться может только 3"-конец.


Сравните нуклеотид для РНК с нуклеотидом для ДНК.
Попробуйте узнать, какой это нуклеотид, в таком представлении:

АТФ - свободный нуклеотид
цАМФ - "закольцованная" молекула АТФ

Схема строения нуклеотида

Обратите внимание на то, что активированный нуклеотид, способный наращивать цепочку ДНК или РНК, имеет "трифосфатный хвостик". Именно этим "энергонасыщенным" хвостиком он может присоединиться к уже имеющейся цепочке растущей нуклеиновой кислоты. Фосфатный хвостик сидит на 5-м атоме углерода, так что это положение углерода уже занято фосфатами и предназнено для прикрепления. К чему же его прикрепить? Только к углероду в положении 3". После прикрепления данный нуклеотид сам станет мишенью дла прикрепления следующего нуклеотида. "Принимающая сторона" предоставляет углерод в положении 3", а "прибывающая сторона" цепляется к нему фосфатным хвостиком, находящимся в положении 5". В целом цепочка растёт со стороны 3".
Наращивание нуклеотидной цепочки ДНК
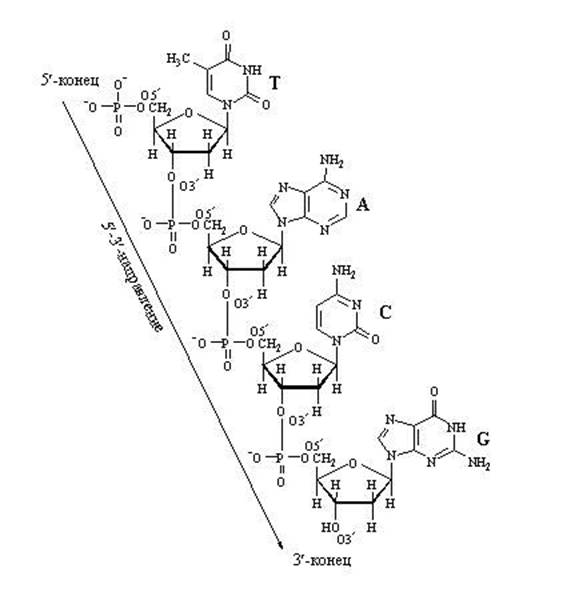
Наращивание цепочки за счёт "продольных" связей между нуклеотидами может идти только в одном направлении: от 5" ⇒ к 3", т.к. новый нуклеотид можно присоединить только к 3"-концу цепочки, но не к 5"-концу.
Пары нуклеотидов, связанные "поперечными" комплементарными связями своих азотистых оснований

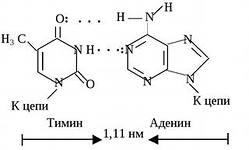
Участок двойной спирали ДНК

Найдите признаки антипараллельности двух цепей ДНК.
Найдите пары нуклеотидов с двойными и тройными комплементарными связями.
Учебное пособие предназначено для студентов направления «Биология» всех профилей подготовки, всех форм обучения для теоретической подготовки к занятиям, зачетам и экзаменам. Пособие охватывает основные разделы структурной биохимии: строение, физико-химические свойства и функции основных классов биологических макромолекул. Большое внимание уделено ряду прикладных аспектов биохимии.
Нуклеотиды и нуклеиновые кислоты
Структура нуклеотидов и азотистых оснований
Нуклеотиды принимают участие во множестве биохимических процессов, а также являются мономерами нуклеиновых кислот. Нуклеиновые кислоты обеспечивают все генетические процессы. Каждый нуклеотид состоит из трех типов химических молекул:
Азотистое основание;
Моносахарид;
1-3 остатка фосфорной кислоты.
В отличие от моносахаридов, нуклеотиды как мономеры являются сложно устроенными молекулами, состоящими из структур, относящихся к разным классам химических веществ, поэтому необходимо рассмотреть свойства и структуру этих компонентов по отдельности.
Азотистые основания
Азотистые основания относятся к гетероциклическим соединениям. В состав гетероцикла помимо атомов углерода входят атомы азота. Все азотистые основания, входящие в нуклеотиды относят к двум классам азотистых оснований: пуриновые и пиримидиновые. Пуриновые основания это производные пурина – гетероцикла, состоящего из двух циклов, один пятичленный, второй – шести, нумерация осуществляется так, как показано на рисунке. Пиримидиновые основания являются производными пиримидина и состоят из одного шестичленного цикла, нумерация также указана на рисунке (Рисунок 31). Главные пиримидиновые основания и у прокариот, и у эукариот – это цитозин, тимин и урацил. Из пуриновых оснований чаще всего встречаются аденин и гуанин. Два других – ксантин и гипоксантин – являются интермедиатами в процессах их метаболизма. У человека в роли конечного продукта катаболизма пуринов выступает окисленное пуриновое основание – мочевая кислота . Помимо пяти названных выше главных оснований известны и менее широко представленные минорные основания. Некоторые из них присутствуют только в нуклеиновых кислотах бактерий и вирусов, но многие также найдены в составе про- и эукариотических ДНК и транспортных и рибосомных РНК. Так, и бактериальная ДНК, и ДНК человека содержат значительные количества 5-метилцитозина; в бактериофагах обнаружен 5-гидроксиметилцитозин. Необычные основания выявлены в матричной РНК – N 6 -метиладенин, N 6 , N 6 -диметиладенин и N 7 -Meтилгуанин. У бактерий также обнаружен модифицированный урацил с присоединенной по N 3 -положению (α-амино, α-карбокси) -пропильной группой. Функции этих замещенных пуринов и пиримидинов до конца не выяснены, однако они могут образовывать неканонические связи между основаниями (это будет рассмотрено ниже), обеспечивая образование вторичных и третичных структур нуклеиновых кислот.
Рисунок 31. Структура азотистых оснований
В клетках растений выявлена серия пуриновых оснований с метильными заместителями. Многие из них фармакологически активны. В качестве примера можно привести кофейные зерна, содержащие кофеин (1,3, 7-триметилксантин), чайный лист, содержащий теофиллин (1, 3-диметил-ксантин), и какао-бобы, в состав которых входит теобромин (3, 7-диметилксантин).
изомерия и Физико-химические свойства пуриновых и пиримидиновых оснований
Молекула азотистого основания образует систему чередующихся одинарных и двойных связей (систему сопряженных двойных связей). Такая организация образует жесткую молекулу, без возможности конформационных переходов. В результате нельзя говорить об изменении конформации азотистых оснований.
Для азотистых оснований выявлен только один тип изомерии кето-енольный переход или таутомерия.
Таутомерия
Благодаря феномену кето-енольной таутомерии нуклеотиды могут существовать либо в лактимной, либо в лактамной формах, причем в физиологических условиях лактамная форма превалирует у гуанина и тимина (Рисунок 32). Важность этого обстоятельства станет ясна при обсуждении процессов спаривания оснований.
Рисунок 32. Таутомерия нуклеотидов
Растворимость
При нейтральном рН наименьшей растворимостью обладает гуанин. Следующим в этом ряду стоит ксантин. Мочевая кислота в форме уратов сравнительно неплохо растворяется при нейтральном рН, но очень плохо растворима в жидкостях с более низкими значениями рН, таких, как моча. Гуанин в моче человека в норме отсутствует, а ксантин и мочевая кислота являются ее обычными компонентами. Последние два пурина часто входят в состав камней мочевого тракта.
Поглощение света
За счет системы сопряженных двойных связей все азотистые основания поглощают в ультрафиолетовой части спектра. Спектр поглощения – график распределения оптической плотности в зависимости от длины волны. Для каждого азотистого основания свой спектр поглощения, по нему можно различить растворы различных азотистых оснований или соединений в состав которых входит азотистое основание (нуклеотиды), но максимум поглощения у всех совпадает при длине волны 260 нм. Это позволяет легко и быстро определять концентрацию как азотистых оснований, так нуклеотидов и нуклеиновых кислот. Спектр поглощения также зависит от рН раствора (Рисунок 33).
Рисунок 33. Спектры поглощения различных азотистых оснований
Функции азотистых оснований
Азотистые основания практически не встречаются в свободном состоянии. Исключение составляют некоторые алкалоиды и мочевая кислота.
Азотистые основания выполняют следующие функции:
Входят в состав нуклеотидов;
Часть алкалоидов – азотистые основания, например, кофеин в кофе или теофелин в чае;
Промежуточные продукты обмена азотистых оснований и нуклеотидов;
Мочевая кислота – причина мочекаменной болезни;
В виде мочевой кислоты выводится азот у некоторых организмов.
Нуклеотиды и нуклеозиды
Молекулы нуклеозидов построены из пуринового или пиримидинового основания, к которому (β-связью присоединен углевод (обычно D-рибоза или 2-дезоксирибоза) в N 9 или N 1 ‒положении соответственно. Таким образом, адениновый рибонуклеозид (аденозин) состоит из аденина и D-рибозы, присоединенной в положении N 9 ; гуанозин – из гуанина и D-рибозы в положении N 9 ; цитидин – из цитозина и рибозы в положении N 1 ; уридин – из урацила и рибозы в положении N 1 . Таким образом в пуриновых нуклеозидах (нуклеотидах) азотистое основание и сахар связаны 1-9 β гликозидной связью, а в пиримидинах – 1-1 β гликозидной связью.
В состав 2́-дезоксирибонуклеозидов входят пуриновые или пиримидиновые основания и 2́-дезоксирибоза, присоединенная по тем же атомам N 1 и N 9 . Присоединение рибозы или 2́-дезоксирибозы к кольцевой структуре основания происходит за счет относительно кислотолабильной N-гликозидной связи (Рисунок 34).
Нуклеотиды – это производные нуклеозидов, фосфорилированные по одной или более гидроксильным группам остатка рибозы (или дезоксирибозы). Так, аденозинмонофосфат (AMФ или аденилат) построен из аденина, рибозы и фосфата. 2́-дезоксиаденозинмонофосфат (дAMФ или дезоксиаденилат) представляет собой молекулу, состоящую из аденина, 2́-дезоксирибозы и фосфата. Обычно к урацилу присоединена рибоза, к тимину – 2́-дезоксирибоза. Поэтому тимидиловая кислота (ТМФ) состоит из тимина, 2́-дезоксирибозы и фосфата. Кроме вышеперечисленных форм нуклеотидов обнаружены и нуклеотиды необычной структуры. Так, в молекуле тРНК выявлен нуклеотид, в котором рибоза присоединяется к урацилу в пятом положении, т. е. не азот-углеродной связью, а углерод-углеродной. Продукт этого необычного присоединения назван псевдоуридином (ψ). Молекулы тРНК содержат и другую необычную нуклеотидную структуру – тимин, соединенный с рибозомонофосфатом. Этот нуклеотид образуется уже после синтеза молекулы тРНК путем метилирования остатка УMФ S-аденозилметионином. Псевдоуридиловая кислота (ψМФ) тоже образуется в результате перегруппировки УMФ после синтеза тРНК.
Рисунок 34. Структура пуриновых и пиримидиновых нуклеозидов и нуклеотидов
Номенклатура, физико-химические свойства и функции нуклеозидов и нуклеотидов
Положение фосфатной группы в молекуле нуклеотида указывается цифрой. Например, аденозин с фосфатной группой, присоединенной к 3-му углероду рибозы, должен быть обозначен как 3́-монофосфат. Штрих после цифры ставят для того, чтобы отличить номер углерода в пуриновом или пиримидиновом основании от положения этого атома в остатке дезоксирибозы. При нумерации атомов углерода основания штрих не ставится. Нуклеотид 2́-дезоксиаденозин с фосфатным остатком при углероде-5 молекулы сахара обозначается как 2́-дезоксиаденозин-5́-монофосфат. Нуклеозиды, содержащие аденин, гуанин, цитозин, тимин и урацил, принято обозначать буквами A, Г, Ц, Т и У соответственно. Наличие буквы d (или д) перед сокращением обозначает, что углеводным компонентом нуклеозида является 2́-дезоксирибоза. Гуанозин, содержащий 2́-дезоксирибозу, может быть обозначен дГ (дезоксигуанозин), а соответствующий ему монофосфат с фосфатной группой, присоединенной к третьему атому углерода дезоксирибозы, – дГ-3́-МФ. Как правило, в тех случаях, когда фосфат присоединен к углероду-5 рибозы или дезоксирибозы, символ 5́ опускается. Так, гуанозин 5́-монофосфат принято обозначать ГМФ, а 5́-монофосфат 2́-дезоксигуанозина сокращают как дГМФ. Если к углеводному остатку нуклеозида присоединены 2 или 3 остатка фосфорной кислоты используются аббревиатуры ДФ (дифосфат) и ТФ (трифосфат). Таким образом, аденозин + трифосфат с тремя фосфатными группами в 5́-положении углевода будет обозначаться АТФ. Поскольку в молекулах нуклеотидов фосфаты находятся в виде ангидридов фосфорной кислоты, т. е. в состоянии с низкой энтропией, их называют макроэргами (обладающими большим запасом потенциальной энергии). При гидролизе 1 моля АТФ до AДФ высвобождается 7,3 кКал потенциальной энергии.
Рисунок 35. Структура цАМФ
Физико-химические свойства нуклеотидов
Так как в состав нуклеотидов входят азотистые основания, то такие свойства как таутомерия и способность поглощать в ультрафиолетовой части спектра также характерны и для нуклеотидов, причем спектры поглощения азотистых оснований и содержащих эти основания нуклеотидов сходны. Наличие сахара и остатков фосфорной кислоты делает их более гидрофильными чем азотистые основания. Все нуклеотиды являются кислотами, так как содержат остатки фосфорной кислоты.
Функции природных нуклеотидов
Нуклеотиды являются мономерами нуклеиновых кислот (РНК, ДНК). В состав ДНК входят дезоксирибонуклеотидфосфаты – производные аденина, тимина, гуанина и цитозина. Также некоторые молекулы гуанина и цитозина в составе ДНК метилированы, то есть содержат метильную группу. Как основные мономеры в состав РНК входят рибонуклеотидфосфаты – производные аденина, урацила, гуанина и цитозина. Также в состав РНК входят нуклеотиды, содержащие различные минорные азотистые основания, например ксантин, гипоксантин, дигидроуридин и др.
Нуклеотиды являются мономерами коферментов (НАД, НАДФ, ФАД, ко-энзим А, метионин-аденозин). В составе коферементов они участвуют в ферментативных реакциях. Более подробно эта функция будет рассмотрена ниже.
Энергетическая (АТФ) . АТФ выполняет функцию основного внутриклеточного переносчика свободной энергии. Концентрация наиболее распространенного свободного нуклеотида в клетках млекопитающих – АТФ – составляет около 1 ммоль/л.
Сигнальная (цГМФ, цАМФ) (Рисунок 35). Циклический AMФ (3́-, 5́-аденозинмонофосфат, цАМФ) – медиатор различных внеклеточных сигналов в клетках животных – образуется из АТФ в результате реакции, катализируемой аденилатциклазой. Активность аденилатциклазы регулируется комплексом взаимодействий, многие из которых инициируются через рецепторы гормонов. Внутриклеточная концентрация цАМФ (около 1 мкмоль/л) на 3 порядка ниже концентрации ATФ. Циклический цГМФ (3́-, 5́-гуанозинмонофосфат, цГМФ) служит внутриклеточным проводником внеклеточных сигналов. В некоторых случаях цГМФ выступает в роли антагониста цАМФ. цГМФ образуется из ГТФ под действием гуанилатциклазы – фермента, имеющего много общего с аденилатциклазой. Гуанилатциклаза, как и аденилатциклаза, регулируется различными эффекторами, в том числе и гормонами. Как и цАМФ, цГМФ гидролизуется фосфодиэстеразой до соответствующего 5́-монофосфата.
Регуляторная (ГТФ) . Активность группы белков (G-белков), выполняющих в основном регуляторную функцию, зависит от того: какой нуклеотид они связывают. В неактивной форме эти белки связывают ГДФ, при активации белка происходит замена ГДФ на ГТФ. При выполнении своей функции белок гидролизует ГТФ до ГДФ и фосфата, выделившаяся, энергия затрачивается на функционирование белка.
Активация при метаболизме липидов и моносахаридов (УТФ, СТФ) . Производные урациловых нуклеотидов участвуют в качестве активирующих агентов в реакциях метаболизма гексоз и полимеризации углеводов, в частности при биосинтезе крахмала и олигосахаридных фрагментов гликопротеинов и протеогликанов. Субстратами в этих реакциях являются уридин-дифосфатсахара. Например, уридиндифосфатглюкоза служит предшественником гликогена. Также превращение глюкозы в галактозу, глюкуроновую кислоту или другие производные моносахаридов происходит в виде коньюгата с УДФ. СТР необходим для биосинтеза некоторых фосфоглицеридов в тканях животных. Реакции с участием церамида и ЦДФ-холина приводят к образованию сфингомиелина и других замещенных сфингозинов.
Участие в дезактивации различных спиртов и фенолов (УДФ-глюкуроновая кислота). Уридиндифосфатглюкуроновая кислота – выполняет функцию «активного» глюкуронида в реакциях конъюгирования, например, при образовании глюкуронида билирубина.
Нуклеотиды в составе коферментов
Коферменты – это низкомолекулярные соединения связанные с ферментами (см раздел «Ферменты») непосредственно участвующие в в биохимической реакции, другими словами это еще один субстрат, не выходящий в окружающую среду.
Коферменты подразделяют на две группы:
переносчики протонов и электронов, эти коферменты участвуют в окислительно-восстановительных реакциях;
переносчики всех остальных групп кроме протонов и электронов, эти коферменты участвуют в трансферазных реакциях.
Более подробно механизмы упомянутых реакций можно рассмотреть в главе «Ферменты».
Некоторые коферменты содержат в своем составе нуклеотиды. Они также делятся на эти же две группы.
Коферменты переносчики протонов и электронов
Эти коферменты участвуют в окислительно-восстановительных реакциях, где аденозин выполняет только структурную функцию, в реакцию вступают нуклеотиды, содержащие другие типы оснований, выделяют два типа таких коферментов: никотиновые и флавиновые. Они отличаются не только по активной группировке, но и по типу реакций, которые они осуществляют.
Никотиновые коферменты
Рисунок 36. Никотиновые коферменты. А-структура NAD, Б-структура NADP, В-механизм активности никотиновой кислоты, Г-механизм работы никотиновых коферментов
Никотинамидадениндинуклеотид (NAD +) – главный акцептор электронов при окислении топливных молекул. Реакционноспособная часть NAD + – его никотинамидное кольцо. При окислении субстрата никотинамидное кольцо NAD + присоединяет ион водорода и два электрона, которые являются эквивалентами гидрид-иона. Восстановленная форма этого переносчика – NADH. В ходе этого дегидрирования один атом водорода субстрата прямо переносится на NAD + , тогда как второй переходит в растворитель. Оба электрона, теряемые субстратом, переносятся на никотинамидное кольцо. Роль донора электронов в большинстве процессов восстановительного биосинтеза (пластического обмена); выполняет восстановленная форма никотин амидадениндинуклеотидфосфата (NADPH). NADPH отличается от NAD наличием фосфата, связанного эфирной связью с 2́-гидроксильной группой аденозина. Окисленная форма NADPH обозначается как NADP + . NADPH переносит электроны таким же образом, как NADH. Однако, NADPH используется почти исключительно в процессах восстановительного биосинтеза, тогда как NADH используется преимущественно для генерирования АТР. Дополнительная фосфатная группа NADPH – это участок, ответственный за осуществление целевого предназначения молекулы, состоящего в распознавании ферментами.
Флавиновые коферменты
Первый флавиновый кофермент (флавинмононуклеотид FMN) был выделен А. Сент-Дьёрдьи из сердечной мышцы в 1932 г., Р. Г. Варбург и В. Христиан тогда же получили из дрожжей первый флавопротеид, содержащий FMN в качестве кофермента. Второй важнейший флавиновый кофермент – флавинадениндинуклеотид (FAD) выделен ими же как кофактор оксидазы D-аминокислот в 1938 году. За счет окислительно-восстановительного превращения флавинового кольца флавиновые коферменты осуществляют окислительно-восстановительные реакции в составе многих важнейших ферментных систем: оксидаз (в частности, оксидаз D- и L-аминокислот, моноаминооксидазы, регулирующей уровень катехоламинов в крови) и дегидрогеназ (часто с участием никотинамидадениндинуклеотида и убихинонов).
Рисунок 37. Флавиновые коферменты. А-структура FAD, Б-механизм активности никотиновой кислоты, В-механизм работы флавиновых коферментов
Второй основной переносчик электронов при окислении топливных молекул – флавинадениндинуклеотид. Сокращения, используемые для обозначения окисленной и восстановленной форм этого переносчика – соответственно FAD и FADH 2 . Реакционноспособная часть FAD – это его изоаллоксазиновое кольцо. FAD, подобно NAD + , присоединяет два электрона. Однако FAD в отличие от NAD + присоединяет оба теряемых субстратом атома водорода.
Конец ознакомительного фрагмента.
В организме человека находится большое количество органических соединений, без которых невозможно представить стабильное течение обменных процессов, поддерживающих жизнедеятельность всех . Одними из таких веществ являются нуклеотиды – это фосфорные эфиры нуклеозидов, которые играют важнейшую роль в передаче информационных данных, а также химических реакциях с выделением внутриклеточной энергии.
Как самостоятельные органические единицы формируют наполнительный состав всех нуклеиновых кислот и большинства коферментов. Рассмотрим более подробно, что такое нуклеозидфосфаты и какую роль они играют в человеческом организме.
Из чего состоит вещество нуклеотид. Оно считается крайне сложным эфиром, относящимся к группе кислот фосфора и нуклеозидов, которые по своим биохимическим свойствам относятся к числу N-гликозидов и содержат гетероциклические фрагменты, связанные с молекулами глюкозы и атомом азота.
В природе наиболее распространенными являются нуклеотиды ДНК.
Кроме этого, еще различают органические вещества с похожими характеристиками строения: рибонуклеотиды, а также дезоксирибонуклеотиды. Все они без исключения являются мономерными молекулами, относящимися к сложным по строению биологическим веществам полимерного типа.
Из них формируется РНК и ДНК всех живых существ, начиная от простейших микроорганизмов и вирусных инфекций, заканчивая человеческим организмом.
Остаток молекулярной структуры фосфора среди нуклеозидфосфатов, образует эфирную связь с двумя, тремя, а в некоторых случаях сразу с пятью гидроксильными группами. Практически все без исключения нуклеотиды относятся к числу эфирных веществ, которые образовались из остатков ортофосфорной кислоты, поэтому их связи устойчивы и не распадаются под воздействием неблагоприятных факторов внутренней и внешней среды.
Обратите внимание! Строение нуклеотидов всегда сложное и основывается на моноэфирах. Последовательность нуклеотидов может меняться под воздействием стрессовых факторов.
Биологическая роль
Влияние нуклеотидов на течение всех процессов в организме живых существ изучают ученые, которые исследуют молекулярное строение внутриклеточного пространства.
Исходя из лабораторных заключений, полученных по итогам многолетней работы ученых различных стран мира, выделяют следующую роль нуклеозидфосфатов:
- универсальный источник жизненной энергии, за счет которой происходит питание клеток и соответственно поддерживается нормальная работа тканей, формирующих внутренние органы, биологические жидкости, эпителиальный покров, сосудистую систему;
- являются транспортировщиками глюкозных мономеров в клетках любого типа (это одна из форм углеводного обмена, когда употребляемый сахар, под воздействием пищеварительных ферментов трансформируется в глюкозу, которая разносится в каждый уголок организма вместе с нуклеозидфосфатами);
- выполняют функцию кофермента (витаминные и минеральные соединения, которые способствуют обеспечению клеток питательными веществами);
- сложные и циклические мононуклеотиды являются биологическими проводниками гормонов, распространяющихся вместе с потоком крови, а также усиливают действие нейронных импульсов;
- аллостерическим образом регулируют активность пищеварительных ферментов, вырабатываемых тканями поджелудочной железы.
Нуклеотиды входят в состав нуклеиновых кислот. Они соединены тремя и пятью связями фосфодиэфирного типа. Генетики и ученые, посвятившие свою жизнь молекулярной биологии, продолжают лабораторные исследования нуклеозидфосфатов, поэтому ежегодно мир узнает еще больше интересного о свойствах нуклеотидов.
 Последовательность нуклеотидов – это разновидность генетического равновесия и баланса расположения аминокислот в структуре ДНК, своеобразный порядок размещения остатков эфира в составе нуклеиновых кислот.
Последовательность нуклеотидов – это разновидность генетического равновесия и баланса расположения аминокислот в структуре ДНК, своеобразный порядок размещения остатков эфира в составе нуклеиновых кислот.
Он определяется с помощью традиционного метода секвенирования отобранного для анализа биологического материала.
Т – тимин;
А – аденин;
G – гуанин;
С – цитозин;
R – GA аденин в комплексе с гуанином и основаниями пурина;
Y – TC пиримидиновые соединения;
K – GT нуклеотиды, содержащие кетогруппу;
M – AC входящие в аминогруппу;
S – GC мощные, отличающиеся тремя водородными соединениями;
W – AT неустойчивые, которые образуют только по две водородные связи.
Последовательность нуклеотидов может меняться, а обозначения латинскими буквами необходимы в тех случаях, когда порядок расположения эфирных соединений неизвестен, является несущественным либо уже имеются результаты первичных исследований.
Наибольшее количество вариантов и комбинаций нуклеозидфосфатов свойственно для ДНК. Для записи эфирных соединений РНК достаточно символов A, С, G, U. Последнее литерное обозначение является веществом уридин, которое встречается только в РНК. Последовательность символических обозначений всегда записывается без использования пробелов.
Полезное видео: нуклеиновые кислоты (ДНК и РНК)
Сколько нуклеотидов в ДНК
Для того, чтобы максимально подробно понимать, о чем идет речь, следует иметь четкое представление о самой ДНК. Это отдельный вид молекул, которые имеют вытянутую форму и состоят из структурных элементов, а именно – нуклеозидфосфатов. Какое количество нуклеотидов в ДНК? Существует 4 вида эфирных соединений данного типа, входящие в состав ДНК. Это аденин, тимин, цитозин и гуанин. Все они формируют единую цепочку, из которой и образовывается молекулярная структура ДНК.
Впервые строение ДНК было расшифровано в далеком 1953 году американскими учеными Френсисом Криком и Джеймсом Уотсоном. В одной молекуле дезоксирибонуклеиновой кислоты содержится по две цепочки нуклеозидфосфатов. Они размещены таким образом, что внешне напоминают спираль, закручивающуюся вокруг своей оси.
Обратите внимание! Количество нуклеотидов в ДНК неизменное и ограничивается только четырьмя видами — данное открытие приблизило человечество к расшифровке полного генетического кода человека.

При этом строение молекулы имеет одну важную особенность. Все нуклеотидные цепочки обладают свойством комплементарности. Это означает, что друг напротив друга размещаются только эфирные соединения определенного вида. Известно, что напротив тимина всегда расположен аденин. Напротив цитозина не может находится никакое другое вещество кроме гуанина. Такие нуклеотидные пары формируют принцип комплементарности и являются неразделимыми.
Масса и длина
 С помощью сложных математических подсчетов и лабораторных исследований, ученым удалось установить точные физико-биологические свойства эфирных соединений, формирующих молекулярную структуру дезоксирибонуклеиновой кислоты.
С помощью сложных математических подсчетов и лабораторных исследований, ученым удалось установить точные физико-биологические свойства эфирных соединений, формирующих молекулярную структуру дезоксирибонуклеиновой кислоты.
Известно, что протяжная длина одного внутриклеточного остатка, состоящего из аминокислот в единой полипептидной цепи – 3,5 ангстрем. Средняя масса одного молекулярного остатка равна 110 а.е.м.
Кроме этого, еще выделяют мономеры нуклеотидного типа, которые сформированы не только из аминокислот, но имеют и эфирные составляющие. Это мономеры ДНК и РНК. Их линейная длина измеряется непосредственно внутри нуклеиновой кислоты и составляет не менее 3,4 ангстрем. Молекулярный вес одного нуклеозидфосфата находится в пределах 345 а.е.м. Это исходные данные, которые используются в практической лабораторной работе, посвященной опытам, генетическим исследованиям и прочей научной деятельности.
Медицинские обозначения
Генетика, как наука, развивалась еще в период, когда не было исследований строения ДНК человека и других живых существ на молекулярном уровне. Поэтому в период домолекулярной генетики нуклеотидные связи обозначались, как наименьший элемент в структуре молекулы ДНК. Как ранее, так и в настоящее время, эфирные вещества данного типа были подвержены . Она могла быть спонтанной или индуцированной, потому для обозначения нуклеозидфосфатов с поврежденной структурой еще используют термин «рекон».
Для определения понятия наступления возможной мутации в азотистых соединениях нуклеотидных связей, применяют термин «мутон». Данные обозначения больше востребованы в лабораторной работе с биологическим материалом. Также используются учеными генетиками, которые изучают устройство молекул ДНК, пути передачи наследственной информации, способы ее шифрования и возможные комбинации генов, получаемых в результате слияния генетического потенциала двух половых партнеров.
Вконтакте